|
|
|
|
Brain Engineering Laboratory: Research
|
|
|
|
Research Part 1: Introduction
Research Part 2: Paleocortical precursors
Research Part 3: Synaptic long term potentiation (LTP)
Research Part 4: Neocortex
Research Part 5: Applications
2. Paleocortical precursors
Modules of the brain. Each telencephalic brain area (cortex, hippocampus, amygdala, striatal complex) is different in structure and operation. Even constituent circuits (fields CA1 vs CA3 of hippocampus; central, medial, lateral divisions of amygdala; layers of cortex) can be dramatically different from each other. Our models of individual brain circuits have yielded quite different operations for each region, corresponding to their different circuit designs. Each emergent algorithm constitutes a candidate function that can be contributed to overall brain computations. Moreover, still other functions arise from interaction of distinct circuits, which combine in sometimes unanticipated ways.
Olfaction as precursor. Early work in our lab focused on the olfactory system. The primary olfactory cortical circuit is substantially simpler than those that process other senses such as sight, sound and touch. Yet the olfactory cortex (aka piriform, pyriform, prapyriformus) bears many resemblances to portions of the other primary cortices, and is considered their direct evolutionary precursor. Thus it may serve as a partial Rosetta stone to aid us in our long-term goal of decoding the (more complex) neocortical areas.
Excitatory-inhibitory interaction. Excitation and inhibition in cortical circuits are very asymmetrical: excitatory cells generate fast (~15 msec) responses and have axons that travel long distances, whereas inhibitory neurons generate slower and longer responses (~80-100 msec), inhibit only cells in a local region, and synapse on or near excitatory cell bodies, strengthening their effect. When inputs arrive at a group of excitatory neurons, their voltages rise until a subset of them responds (to a sufficiently strong input), at which point their activity triggers local feedback inhibition, which quiets all the cells in the inhibitory cells' local region, including the cells that gave rise to the inhibiton. The result is a "race" in which the most strongly activated neurons respond, and other neurons are prevented from responding via feedback inhibition. This "winners take all" effect has been described and formalized [Coultrip, R., Granger, R., and Lynch, G. (1992). A cortical model of winner-take-all competition via lateral inhibition. Neural Networks, 5: 47-54.]
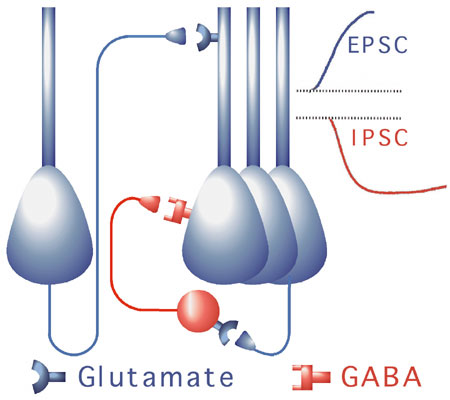
Simulation of Paleocortex. The primary characteristics of olfactory paleocortex that have been modeled are the feedforward connections from bulb to cortex, and feedback connections from cortex back to inhibitory (granule) cells in bulb, along with the local cortical excitatory-inhibitory interaction just described.
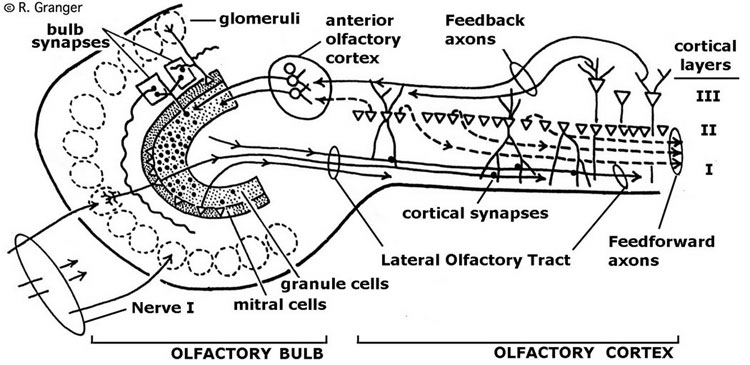
Operation of the simulation. The paleocortical model, like the biological system, responds repeatedly to a stimulus, eliciting rhythmic responses roughly every 200 msec, or five times per second (the "theta" EEG rhythm, which is strongly in evidence in animals actively engaged in olfactory behavior). Surprisingly, the model generated a different response at each such cycle, so that when presented with a simulated odor, it generates a series of distinct cortical response patterns. Analysis showed that with learning via simulated long-term potentiation (LTP), these responses corresponded to a series of general to specific categories of the input. The result was compared to standard statistical methods for hierarchical clustering, and was found to have time and space costs that compared favorably with comparable algorithms in this class. [Ambros-Ingerson, J., Granger, R., and Lynch, G. (1990). Simulation of paleocortex performs hierarchical clustering. Science, 247: 1344-1348.]
Simulation of Paleocortex Performs Hierarchical Clustering. Operation of the olfactory simulation creates memories organized hierarchically, which are traversed or read out in sequence, from general to specific. Thus, for demonstration purposes, the simulation was presented with a series of brief speech sounds: spoken letters of the alphabet. Once they were learned by the system, then presentation of any letter would evoke a sequence of responses. For instance, presenting "B" to the system elicited an initial response corresponding to a spontaneously created category containing all "B", "C", "D", "E", "G" sounds, followed by a response corresponding only to the sub-category continaing unvoiced "B", "D", "E"; followed by a response specific just to "B". Presenting "C" to the system elicited the same initial response, followed by a response corresponding to the sub-category of voiced "C", "G" sounds, followed by a response specific just to "C". [Granger, R. (2002). Neural Computation: Olfactory cortex as a model for telencephalic processing. Learning & Memory (J.Byrne, Ed.), pp.445-450.]
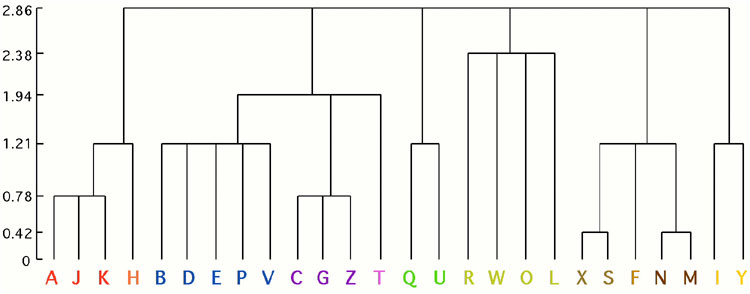
Experimental tests of olfactory paleocortical model.
Behavioral tests of the simulation investigated whether animals would form spontaneous clusters of similar odors, as the model does. Animals were presented with a number of similar odors, and then tested on novel odors that were also similar to the ones trained on. In a series of experiments, it was shown that the animals treated the novel odor as though it were recognized, and yet could clearly distinguish between the novel odor vs the familiar ones [Granger, R., Staubli, U., Powers, H., Otto, T., Ambros-Ingerson, J., and Lynch, G. (1991). Behavioral tests of a prediction from a cortical network simulation. Psychol. Sci., 2: 116-118].
Physiological predictions of the model were tested via recording electrodes implanted into the paleocortex of rats who then participated in a series of olfactory learning experiments. When the animals learned many new odors, cells in cortex selectively responded with to specific odors, with brief burst responses occurring time-specifically at about 200 msec after onset of the odor. These responses contrast sharply with those obtained from animals who are not made familiar with the task, are not actively engaged in rapidly learning many distinct odors, or who are not freely moving during the experiment [McCollum, J., Larson, J., Otto, T., Schottler, F. Granger, R., and Lynch, G. (1991). Short-latency single-unit processing in olfactory cortex. J. Cog. Neurosci., 3: 293-299]
Hippocampal models. Just as the paleocortex is a precursor of neocortex, the hippocampal archicortex has many characteristics of cortex, including laminar organization and glutamatergic synapses exhibiting LTP. The dentate gyrus and field CA3 contain many ancient characteristics distinguishing them from cortex (including non-pyramidal excitatory cells (granule cells) in dentate, mossy fiber synapses (anatomically and physiologically not at all like cortical synapses, and not exhibiting cortical LTP), and extensive excitatory feedback axon collaterals in CA3, unlike most cortical arrangements. Field CA1 and subiculum, however, share many characteristics with cortex. Models of these fields led to identification of a mechanism for time dilation in CA3, in which an input from entorhinal and dentate were maintained via recurrent activity in CA3 axon collaterals until subsequent input arrived, allowing arbitrary delay latencies between pairs of inputs, when integrated into a model of the three primary components of hippocampus [Granger, R., Wiebe, S., Taketani, M., Ambros-Ingerson, J., Lynch, G. (1997). Distinct memory circuits comprising the hippocampal region. Hippocampus, 6: 567-578].
|
© 2005 TSG.RHG
[GrangerLab]
|
|
