|
|
|
|
Brain Engineering Laboratory: Research
|
|
|
|
Research Part 1: Introduction
Research Part 2: Paleocortical precursors
Research Part 3: Synaptic long term potentiation (LTP)
Research Part 4: Neocortex
Research Part 5: Applications
4. Neocortex
Neocortex and the thalamocortical system. Based on these serial studies of paleocortex, hippocampus cortical local circuits, and the physiology and biochemistry of synaptic change via LTP, initial forays were made in the late nineties, into the most important systems of the brain: thalamocortical circuits. The neocortex, which makes up the vast majority of human brain, has a more complex structure than either paleocortex or hippocampus, and is reciprocally linked to subregions of the thalamus via large feedforward (thalamus to cortex) and feedback (cortex to thalamus) pathways of axons.
(A note on quantitative neuroanatomy. "Parallel computers," in which many computing elements are wired together to operate in parallel, offer a modest analogy with the brain. If you look inside a parallel computer, you will see mostly just wires. This is because for every computing element added, it is typically necessary to add wires from it to all the other computers. The number of wires thus grows much faster than the number of computers: 3 wires for 3 computers, 6 wires for 4 computers, 10 wires for 5 computers, 15 wires for 6 computers, etc. In the brain, with billions of neurons (computing elements), wiring them all together would require a brain the size of a football field. Instead, the brain only selectively and sparsely wires neurons together, omitting any wires (axons) that it can in order to enable it to function. This sparse connectivity leads to quite different mathematical operation of the system than if it were completely (densely) wired. It also give us a hint of sorts -- as mammals grow larger with evolution, and the brain grows correspondingly larger, the "cost" of wires grows severe. Even though neuroanatomists sometimes find evidence of axons connecting many different brain areas, they also find quantitatively that the number of axons differ considerably, with some areas heavily linked to others by large axon tracts and others much less strongly linked. In large brained mammals (such as humans), anywhere large tracts of axons appear, this is widely seen by neuroscientists as evidence of strong relative importance of the pathways, in light of the intense pressure to cut back on any less-than-necessary axon tracts. Thus, in our models, we emphasize large pathways and attend less to smaller pathways, aware that something will be missed, but hoping (calculating) that the more essential elements will this way be represented.)
Anatomical structure of the thalamocortical system. The structure of the system is outlined in the figure below. Since not everything can or should be represented in a simulation, and for the evolutionary reasons just described, large axon pathways are emphasized at the expense of smaller ones.
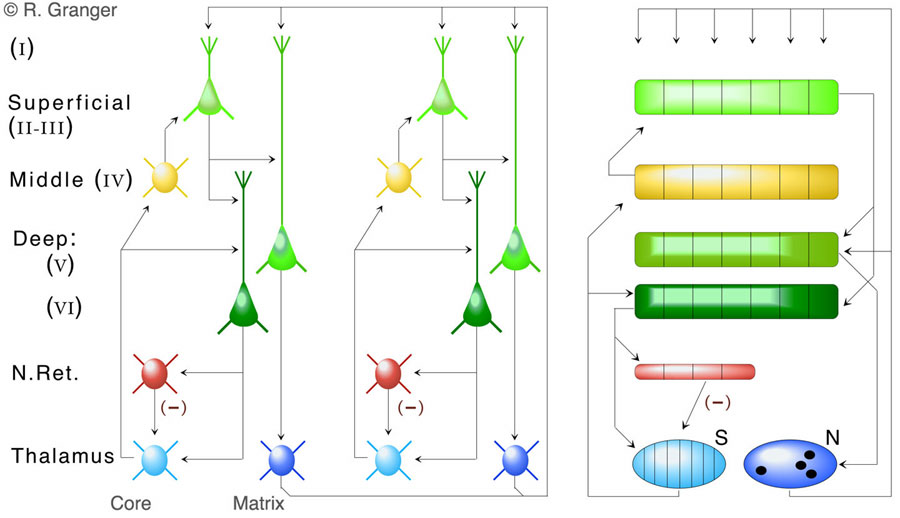
Thalamocortical modeling: Static inputs. Neocortex (e.g., auditory cortex) can receive streams of inputs that are rapidly changing over time, but can also pay sustained attention to a static or slowly-changing input (e.g., a hum, an image). A model of the superficial layers of neocortex found that, when presented with static inputs, an effect related to that in paleocortex occurred: inhibitory feedback from cortex via the inhibitory thalamic reticular nucleus to the thalamic inputs to cortex effectively inhibited or masked the part of the input that cortex had responded to, and thus sustained attention to an input yielded a series of different responses from cortex, corresponding to hierarchical information (category, subcategory, etc). This finding generates specific predictions for human recognition performance. We have tested these in a series of perception experiments on human subjects, with findings supporting the predictions: there is evidence that spontaneous unsupervised categories are constructed, that they are accessed in ascending order of specificity (category, subcategory, etc), and that each step in this serially-performed recognition takes a fixed quanta of time (about 150 msec). These new experimental findings have been written up and submitted for publication [Granger, R., Kilborn, K., Rodriguez, A., and Cabanne, C. Evidence that categorization and recognition are performed by a single thalamocortical mechanism. (In submission)].

Thalamocortical modeling: Time-varying inputs. Further modeling efforts have focused on the processing of superficial and deep neocortical layers in response to changing input streams such as speech or music. Superficial layers are initially activated as described above, but since subsequent inputs are not the same as previous ones (i.e., the input is not static), the input elicits a sequence of different superficial layer responses. Deep layers (V and VI) receive brief sequences of activation from superficial layer and from thalamic "matrix," or nonspecific thalamic nuclei (which strongly innervate all areas of neocortex). The non-topographic connectivity of thalamic matrix nuclei yield a broad range of different cortical responses. The result is a model that produces quite different deep layer encodings for even very similar inputs, i.e., the cortical encodings orthogonalize or emphasize the differences among inputs. Analyses have demonstrated the relationship between this function and that of the common computational algorithm of "hash" coding, which is widely used for efficient storage of large bodies of data. Thus we have forwarded the hypothesis that the operation of cortical deep layers contributes in a specific way to a very high-capacity memory storage mechanism in the brain. [Rodriguez, A., Whitson, J., and Granger, R. (2004) Derivation and analysis of basic computational operations of thalamocortical circuits. Journal of Cognitive Neuroscience, 16:856-877.].
The figure shows the probability that an error of retrieval will occur, for a given number of input sequences stored in a relatively large thalamocortical model. The network consists of 26 cortical "columns" or modules, each containing 100,000 synapses on deep layer neurons. The inputs to the simulation are sequences of 20 letters, chosen from the alphabet of 26 letters, each corresponding to one of the simulated "columns." The curves show the mathematically calculated number of errors that should occur as the number of 20-letter sequences stored increases. Curves are shown for conditions when 4 synapses versus 5 synapses are potentiated for each input letter (n=4 vs n=5). The three points shown are the number of actual errors that occurred in a simulation containing 100,000 synapses, after attempts to store 50,000, 120,000, and 160,000 20-letter sequences, respectively.

Audition, vision, association. Early mammalian brains were dominated by olfaction and by the new neocortical structures that had just evolved. These new structures were represented largely in the new auditory system: although audition, vision, and touch cortex were similar, it is hypothesized that the auditory system was the prototype for the nocturnal therapsids, or early mammals. Although unique specializations occur in the primary sensory cortical areas and elsewhere, most of neocortex is remarkably similar throughout the brain. The models described here are intended to correspond to the shared generic characteristics of most of neocortex, rather than to any particular specializations of sensory areas. The primary difference between larger and smaller brained mammals is expansion of the neocortical area, with little evidence for new specializations occurring that alter the basic design of each neocortical region. Thus animals with bigger brains have quantitatively more of essentially the same neocortex, rather than qualitatively different structures in their brains. One of the key issues in the modeling of cortex is to capture how it is possible that qualitative differences (e.g., between rats, cats, apes, and humans) arise from what appear to be almost entirely quantitative changes alone.
|
© 2005 TSG.RHG
[GrangerLab]
|
|
